Short Kings, Tall Queens, and the Tribulations of Mayflower
In 1918, a law was passed in Massachusetts making Epigaea repens (mayflower) the state flower. Massachusetts General Laws Part 1, Title 1, Ch. 2, § 7 provides that: “Any person who pulls up or digs up the plant of the mayflower or any part thereof… shall be punished by a fine of not more than fifty dollars; but if a person does any of the aforesaid acts while in disguise or secretly in the nighttime he shall be punished by a fine of not more than one hundred dollars.” This law is on the books, still phrased as it was on May 1, 1918—a fitting date. The tribulations of the mayflower arose from people digging it up to make wreaths or bouquets. This led to local population decline, and the law aimed to thwart that. But the law is not the only intriguing historical aspect of this plant. The plant’s flowers reveal a secret of their evolutionary history.

Epigaea repens flowers blooming along the Cumberland Plateau, Tennessee.
Our species is fascinated by the way flowering plants reproduce. Perhaps it is their ruffled petals, extravagant colors, or alluring perfume that remind us of our own courtship rituals. Or maybe it is the highly curious mode of coercing species from different kingdoms of life to move their gametes around. From the poetry of Emily Dickinson to the paintings of Georgia O'Keeffe, we often associate flowers with sex. Indeed, even Carl Linnaeus’ approach for classifying plants was a sexual system, based on grouping plants according to their number of reproductive bits.
The comparison of flowers to sex has historically made some people uncomfortable. In an essay published by Iowa State University’s library archivist, Amy Bishop explained “many Englishmen of the time were scandalized by the sexual nature of Linneaus’ taxonomic system.” But not everyone was repulsed by this system. Charles Darwin’s grandfather, Erasmus Darwin enjoyed it. His poetry “The Loves of the Plants,” Part II of The Botanic Garden, toes the line between plant science communication and a modern romance novel. On page 31–32, Erasmus writes “Woo'd with long care, Curcuma cold and shy Meets her fond husband with averted eye: Four beardless youths the obdurate beauty move With soft attentions of Platonic love.” His care for plants and his desire to elevate them in the popular consciousness is palpable. But, Erasmus did not just write sensually about plants for the sake of passion, he also used his art to popularize the Linnaean system of botany.
What is fascinating about Erasmus’s work is the juxtaposition between his flowery writing, with the depth of his footnotes; each beautiful poem is accompanied by academic text. The poem of four beardless youths of Curcuma (turmeric) is followed by, “One male and one female inhabit this flower; but there are besides four imperfect males, or filaments, without anthers upon them, called by Linnaeus eunuchs.” Here, Erasmus describes the morphology of turmeric flowers, with only one fertile stamen (his “male”, the structures that produce pollen) and several sterile stamens called staminodes (his eunuchs). This morphology is consistent across the turmeric and ginger family (Zingiberaceae). This is science communication. It beautifully ties the art of poetry with detailed biology.

Plates of floral diagrams from Darwin, Erasmus. The Botanic Garden. Part II: Containing The Love of the Plants. A Poem with Philosophical Notes. 4th ed., J. Moore, 1796.
There is a good reason why flowers are thought to be sexual: this is where it happens. The flower is composed of several distinct parts, each with a usual function. The petals are attractive, usually enticing animal pollinators. The stamens are composed of a stalk (filament) and the anther (full of pollen). The pistil has three distinct parts: the ovary or fruit (where the seeds are housed); the stigma where pollen is deposited; and the style that connects the stigma and the ovary. The style is a critical junction, as it acts as the highway for sperm inside the pollen to get to the ovules.
When pollen lands on the stigma it actually germinates like a seedling, sending a long tube down into the tissue of the style. As the pollen tube grows it physically burrows into the living tissue of the style, and even secretes enzymes to digest the style tissue. A macabre path to fertilization. In the style, each pollen grain may compete with dozens or hundreds of other pollen grains, fighting for a chance to fertilize an ovule. The length of the style can act as a maternal fitness test for pollen grains; a marathon of sorts, where the winner will reproduce.

Flower Diagram. Original diagram from Pearson Scott Foresman. Public Domain
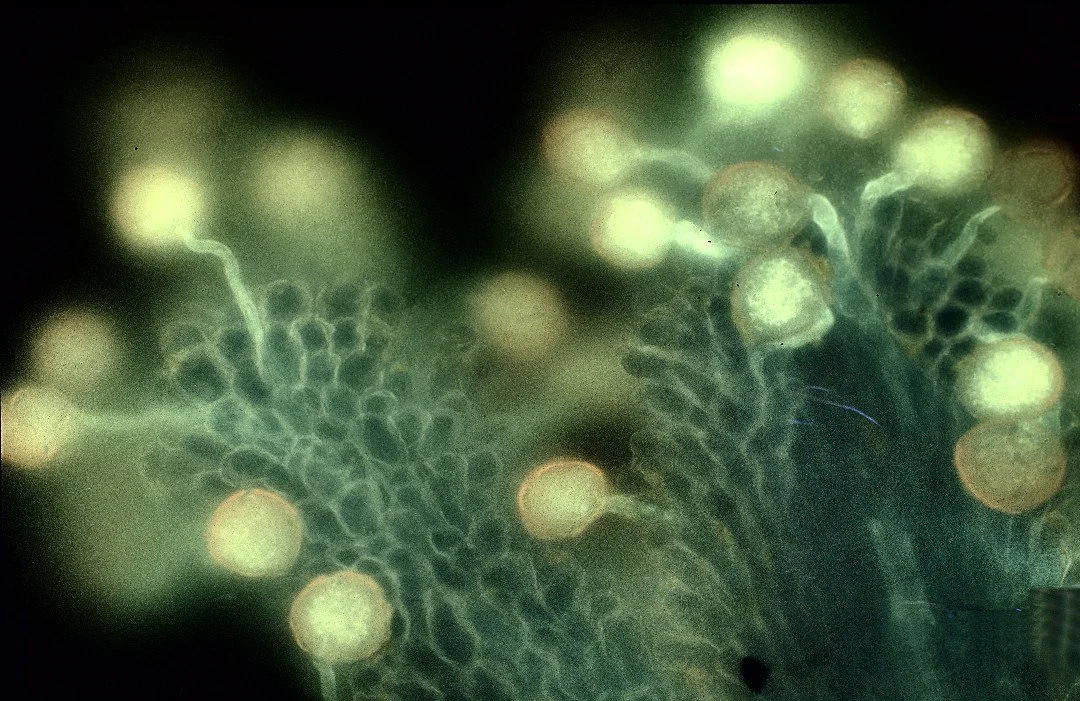
Micrographs showing pollen grains germinating on the stigma of Fameflowers (Talinum). Image by George Shepherd, CC BY-NC-SA 2.0

The position of the style is not only important for germinating pollen and fertilization, but is critical for proper pollination to occur. When a bee or other animal visits a flower they will be dusted with pollen. Where that pollen collects on their body is not necessarily a random process, but one that often co-evolves as the relationship between pollinator and plant becomes more intimate. Some animals are dusted with pollen on their heads, others their backs or abdomens. Upon visiting another flower of the same species, the pollen will then be deposited on the tip of the stigma. Thus, the placement of pollen on the pollinator and the orientation of the stigma will often correspond.
This past month, while I was botanizing in the Cumberland Plateau in Tennessee I closely observed the flowers of E. repens and noticed that different individuals in a single population contained flowers of different types. Some flowers had their style sticking up out of the flower while in other flowers the style was tucked away deep in the floral tube. This seemingly subtle difference has some profound implications for the reproduction and evolutionary dynamics of mayflower. I apologize in advance for the technicality of this essay, but I felt it appropriate if we are to appreciate the full evolutionary and ecological importance of these observations.

What I believe to be pistilate (“female”) flowers, see large stigma sticking through the center. Image by Ben Goulet-Scott.

What I believe to be staminate (“male”) flowers, see pollen grains scattered in the center.
Flowers are usually depicted as having both the stamen (pollen producing structures) and stigma (pollen receiving structures) near each other in what is traditionally called a “perfect” flower. However, this configuration of reproductive parts increases the probability of self-fertilization, or “selfing” (pollen of one flower fertilizing the ovules of the same flower or plant). This would lead to inbreeding. It is conventional knowledge that inbreeding is not ideal and generally, natural selection favors outcrossing. We are reminded of this in royal bloodlines: the health issues of Charles II of Spain were likely a direct product of the consummation between Philip IV and Philip’s niece Mariana of Austria.
Many flowering plants have evolved distinct strategies to inhibit self-fertilization and to promote outcrossing. One of the most well studied mechanisms for avoiding selfing is heterostyly, the phenomenon whereby some individuals in a population will produce flowers that have short styles and other individuals will produce flowers with long styles. Charles Darwin actually wrote about heterostyly in his book The Different Forms of Flowers on Plants of the Same Species (Darwin and Darwin, 1877). He documented this phenomenon in several species, most iconically, primrose (Primula). Philip Gilmartin (2015) elegantly tracked the history of observations of heterostyly and found that these observations date back well before Darwin’s own observations, but in classic fashion, Darwin was able to synthesize the importance behind this observation. It is fitting that Darwin wrote about flowers, maybe his grandfather’s science communication influenced him.
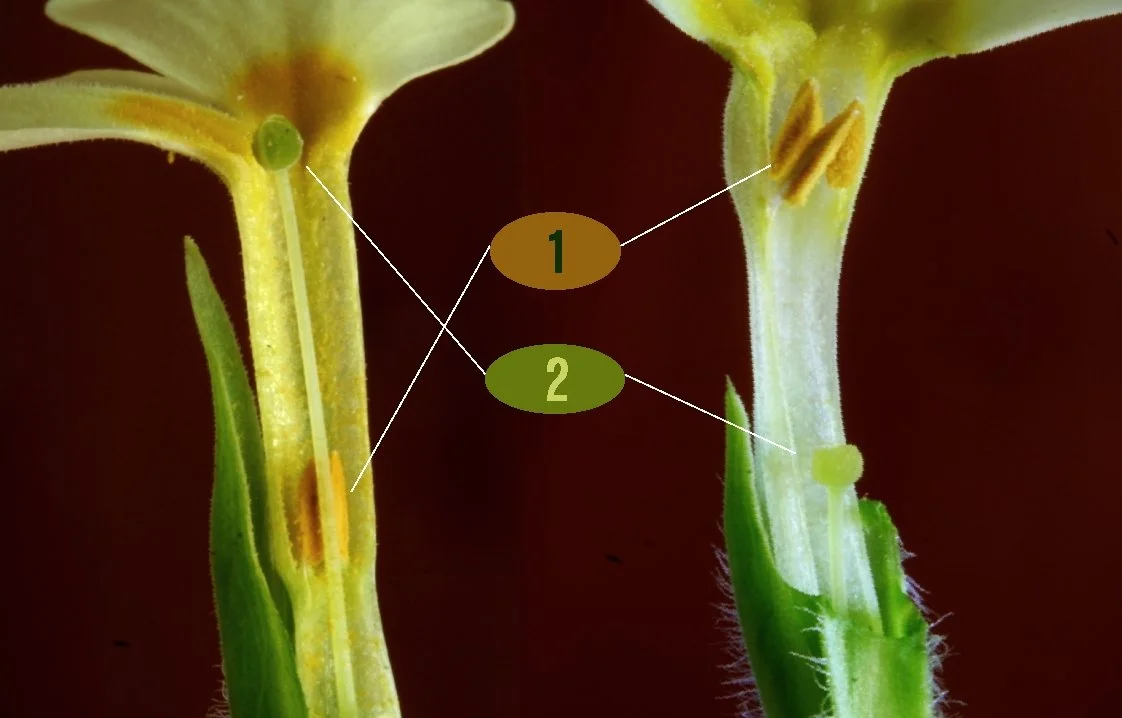
Heterostyly of Primula vulgaris. 1: stamen and 2: style. Image by Enrico Blasutto, CC BY-SA 3.0 (and GFDL).
Darwin proposed that heterostyly promotes outcrossing by reciprocally placing floral organs at different positions. Short-styled flowers are called “thrums” and long-styled flowers are called “pins.” The spatial separation of parts would reduce pollen transfer within the same flower morph—as pollen from one morph is unlikely to reach the stigma of that same morph. Rather, the pollen deposited by long flowers is more likely to reach the stigma of a short flower, and vice versa. As a result, cross-pollination between morphs is favored, rather than within morphs.
But this raises a question: if long and short flower morphs continuously reproduce, how do they maintain their distinctions? Why do they not produce offspring that merge into a continuum of flower morphs, from short to tall? When organisms undergo sexual reproduction they first have to produce their gametes. To do so, they undergo meiosis—the process of cell division which eventually leads to the production of sperm and egg. Meiosis is brilliant. During this critical phase the chromosomes will intertwine and mix with each other, leading to individual sperm and egg cells that are genetically distinct (e.g., each of your sperm or egg cells is slightly different from another one because of this splicing of genes during meiosis). This is why siblings are not 100% identical. So, if there were several genes that were involved in heterostyly, would we not expect that genes responsible for flower size would be recombined, leading to offspring with a slew of different flower morphs all of intermediate size?
In heterostylous plants, the short and long flower morphs are actually determined by several genes clustered into one region of the genome—together called a supergene. The curious point about this supergene is that, instead of having two different types (a short and long form) many heterostylous plants are actually hemizygous, meaning there is only a single version of the gene. Put simply, an individual either has it or does not have it. This means that when individuals reproduce, their offspring will not be a blend of intermediate forms, but rather distinct entities with the supergene (short-styled) or without it (long-styled).
Interestingly, this hemizygosity has independently evolved in several lineages including buckwheat (Fagopyrum esculentum), primrose (Primula vulgaris), and flax (Linum tenue) and other lineages. This represents a convergently evolved mechanism—similar to how bats, birds, and insects all evolved wings. Convergent evolution usually does not land on a nearly identical mechanism—for instance an insect wing is built differently from a bird’s. However, in the case of heterostyly, it seems to have converged on essentially the same mechanism of hemizygosity of a large supergene.
There is a dizzying amount of variation around the theme of heterostyly. Indeed some heterostylous plants do not just differ in the vertical dimensions, but some are mirror images of each other—like chemical enantiomers. The primary reason for all this variation is to ensure outcrossing. If this is the goal, why produce flowers that even have the potential to make pollen and seeds on the same individual? Why not evolve a strategy, like many animals, to produce whole individuals that are male and others that are female? This, too, has evolved in plants and it's called dioecy: the phenomenon whereby a species produces some individuals that only make pollen-bearing flowers (“males”) and others that only produce seed-bearing flowers (“females”). While the exact routes to the evolution of dioecy are unclear, heterostyly could be one intermediate step in its evolution (e.g., first evolve a genetic pathway to flower dimorphism, then to full-on unisexual individuals).
This brings us back to the mayflower. The floral morphology of Epigaea repens has caused confusion since the late 19th century. This plant is actually not heterostylous, as I originally thought. In an 1893 description, W.P. Wilson describes the floral morphology. Before expounding on the biology, he first reminds us of the trials of the mayflower: “Owing to its early appearance, its often beautifully tinted and sweet scented flowers, and its trailing evergreen leaves it is a great favorite wherever it grows… As soon as it opens in the spring it is brought into Boston, tied up into little bunches, with most of its attractive leaves stripped off, and exposed for sale on the streets.” Wilson goes on to explain that there are essentially male and female plants, although the flower morphology is variable between the different sexes. More recent work from Clay and Elistrand (1981) indicate that the mayflower is clearly dioecious, but may have never even been heterostylous. To date, this is the last in-depth work on mayflower floral morphology and it leaves us with many more questions than answers. I will say, I am in good company for my initial misunderstanding of floral morphology in mayflower; Darwin warned about the deception of heterostyly, “Again, a species tending to become dioecious, with the stamens reduced in some individuals and with the pistils in others, often presents a deceptive appearance.” (Darwin and Darwin, 1877, p. 3).
Our own biology tricks us into thinking very narrowly about the possible ways organisms can reproduce. Plants, with their fundamentally distinct processes show us that there are a multitude of different ways to ensure outcrossing and proper fertilization. These small processes of moving reproductive parts a few centimeters higher or lower, or making plants with separate organs, can have profound effects on the way a plant reproduces and its evolutionary fate. Flowers are still for sex; it's just a bit more complex than we would expect.
Further readings and references:
Alvarez, Gonzalo, Francisco C. Ceballos, and Celsa Quinteiro. "The role of inbreeding in the extinction of a European royal dynasty." PloS one 4.4 (2009): e5174.
Barrett, Spencer CH. "‘A most complex marriage arrangement’: recent advances on heterostyly and unresolved questions." New Phytologist 224.3 (2019): 1051-1067.
Bishop, Amy. “Rare Book Highlights: Plants, Sex, and Poetry with Erasmus Darwin.” Cardinal Tales: Highlights from 2018, Iowa State University, 2019, https://iastate.pressbooks.pub/cardinaltales1/chapter/rare-book-highlights-plants-sex-and-poetry-with-erasmus-darwin/
Clay, Keith, and N. C. Ellstrand. "Stylar polymorphism in Epigaea repens, a dioecious species." Bulletin of the Torrey Botanical Club (1981): 305-310.
Darwin, Charles, and Francis Darwin. The different forms of flowers on plants of the same species. Appleton, 1877.
Darwin, Erasmus. The Botanic Garden. Part II: Containing The Love of the Plants. A Poem with Philosophical Notes. 4th ed., J. Moore, 1796.
Gilmartin, Philip M. "On the origins of observations of heterostyly in Primula." New Phytologist 208.1 (2015): 39-51.
Jesson, Linley K., and Spencer CH Barrett. "Solving the puzzle of mirror-image flowers." Nature 417.6890 (2002): 707-707.
Mayer, S. S., and D. Charlesworth. "Cryptic dioecy in flowering plants." Trends in Ecology & Evolution 6.10 (1991): 320-325.
Simón-Porcar, Violeta, et al. "Convergent evolutionary patterns of heterostyly across angiosperms support the pollination-precision hypothesis." Nature Communications 15.1 (2024): 1237.
Wilson, William Powell. "Observations on Epigaea repens L." Contributions from the Botanical Laboratory and the Morris Arboretum of the University of Pennsylvania 1 (1893): 56-63.
Edited by Ben Goulet-Scott